|
今天的科学技术可使人类惬意地遨游太空并骄傲地漫步月球,但面对生命如何起源这一问题却一筹莫展。这毫无疑问将阻碍我们对诸多生命科学现象或问题的认知(如生命的本质和进化,等等)。迄今为止,科学家也提出了各式各样关于生命起源的理论或假设,它们看似玄妙莫测、缤纷瑰丽,但却如流星般地消隐,因为没有一个能经得起绵密的推敲……
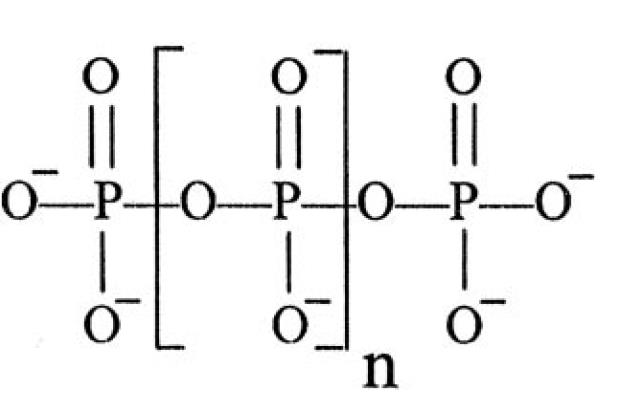
生命源自何方(或生命是如何起源的)?这一问题看似简单,但时至今日人类对它依然知之甚少,或者说,虽然进行了不懈的努力,但是远未获得令人信服的答案。在这里不讨论上帝创世论,因为这不是科学而是一种宗教,也不讨论宇宙生命论,因为,这一学说主张一切生命来自宇宙,但问题是宇宙中的生命又是如何产生的呢?因此,这里限于介绍关于发生在地球表面上产生原始生命的化学进化途径相关的各种假说(关于RNA世界假说,笔者将在下一个博文中单独论述)及笔者自己的见解。我认为,每一种假说,都可能是在揭示真理的一个侧面或一个点,但没有哪一个能画出一个草图(哪怕十分粗犷),或者给我的感觉犹如盲人摸象。 1.“原始汤”假说("Soup" hypothesis) 生命起源于原始地球上有机物的“原始汤(primeval soup)”的思想可追溯至英国博物学家达尔文(CharlesRobert Darwin)、苏联生物化学家奥巴林(Alexander Ivanovich Oparin)等,但是,实验性的证据直到20世纪中期才开始出现。1952年,美国化学家米勒(Stanley Lloyd Miller)模拟闪电的条件,在常温下,用一些无机分子,成功合成了许多有机分子(如氨基酸)。他的实验支持了奥巴林在20世纪初提出的关于生命起源的一些猜想,即在早期的地球上,可能存在一个还原性的原始大气圈(主要是CH4、NH3、H2O和H2等)以及原始的海洋,闪电作用将这些气体聚合成多种氨基酸,即使在常温常压下,这些氨基酸也有可能在局部浓缩,再进一步演化成蛋白质,而蛋白质和其它的多糖类与高分子脂类,在一定的时候就有可能孕育出生命。 已故的美国化学家RobertShapiro(1935–2011)在 “Origins: ASkeptic's Guide to the Creation of Life on Earth”一书中对原始汤假说进行了总结:①早期的大气环境是还原性的,②还原性大气在暴露在各种能量的情况下产生了简单的小分子化合物,③这些化合物浓缩成“汤”,这可能发生在不同的地方(海滨、海洋热液喷口等),④通过进一步的转化,更为复杂的有机聚合物乃至生命在这种有机汤中就发展起来了。①—③已通过实验得到了证实,问题是④如何成为现实仍然是一个迷团(Shapiro 1987)。 笔者认为,米勒及其后续类似的实验研究能够解释在给予足够能量的条件下,从简单的有机物可以生成像氨基酸这样相对简单的有机化合物(虽然这些生成的氨基酸在手性上与蛋白质氨基酸不同)。这只是生命起源迈出的最初一步。 2. 粘土假说(Clay hypothesis) 20世纪60年代中期开始,英国物理化学家凯恩斯•史密斯(Cairns-Smith A. G.)致力于一种称为粘土假说的生命起源学说,他在“Genetic Takeover: and the Mineral Origins of Life”一书中指出,粘土晶体在溶液中的自我复制可能在非生物活性物质和有机生命之间提供了一个简单的中间步骤。粘土在硅酸盐溶液中自然形成,与其它晶体一样,粘土晶体在它们的生长—断裂—再生长过程中,保存其外部的排列形式。一个特定外部形式的粘土晶体可能以一种影响它们进一步复制机会那样的方式去影响其外部环境(Cairns-Smith 1987)。 凯恩斯•史密斯认为,对在其表面捕获一定形式的分子的粘土晶体也存在一种自然选择过程,有利于促进其潜在的复制能力。相当复杂的原有机分子(proto-organic molecules)能被硅酸盐表面特性晶体化,最终,当这些复杂的分子从它们的晶体模子中形成“遗传接管”(genetic takeover)后,就变成了一个独立的复制核心,在进化上的这一刻就是所谓的最初的联适应(the first exaptation)。他将此作为DNA形成的一种可能进化模式。 关于最初的有机分子是如何获得的问题,凯恩斯•史密斯(1995)《生命起源的七条线索》一书中认为是某些矿物质模拟光合作用获得的,“在原始地球上发生的反应不会像现代植物的光合作用那样高深复杂……有些矿物质却可以在有限的程度上模仿这种植物的光合作用。在紫外线的照射下,某些结构简单的铁盐溶于水中之后可以使二氧化碳凝固并将它合成如甲酸之类的小有机分子。某些晶体矿物质也能以这种方式模仿光合作用效应……固氮也是有机物中一种非常复杂的反应过程。这种复杂的反应过程同样也有一些矿物质可在有限程度上加以模仿。作为砂的一种次级成分,含铁的二氧化钛也具有固氮能力。当太阳光照射到这种物质的潮湿的晶体之上,这时就会有少量的氮转化为氨。氨是一种形态的氮,它相比之下比较容易发展成为如氨基酸之类的较大的分子”。 凯恩斯•史密斯(1995)认为,这些小有机分子的“合成当然绝不可能在汪洋大海这样广阔的范围内进行,但在局部,例如说,在潮湿的矿物质暴露在大气层以及早期含有大量紫外线的太阳光线之下时,这种合成是完全可以想象得到的”。 粘土矿物制造这些有机分子对其自身又有何作用呢?凯恩斯•史密斯(1995)说,“甲酸是最简单且最易合成的有机分子之一,它可以稳定溶液的酸性,从而帮助控制粘土结晶过程。比甲酸稍许复杂一些的草酸也相当容易合成。大家知道,草酸能帮助粘土合成……有几种分子通过将铝离子溶解协助粘土合成。氨基酸以及小串的氨基酸,还有其它一些分子,则善于控制溶液中的金属离子……核苷酸的字母段是一种易于粘结在粘土层之间的分子……核苷酸中出现的三价磷酸盐单位则特别适合与粘土层的边缘粘结。也许核苷酸这种分子首先就是为了与粘土相互作用而设计的……多糖由许多连在一起的糖单位合成……当粘土有机物的生长必须解决溶液不因受到光线照射而被烘干或者被水冲掉等诸如此类的问题时,多糖的用途就会明确地显示出来。也许脱氧核糖核酸和核糖核酸的前身属于一种特异多糖。无论是不是,你都可以这样理解:即脱氧核糖核酸或核糖核酸的前身在开始时并不具有遗传用途。发现这类分子可以复制信息是后来的事情,而且这种发现是一种偶然事件……某种东西为了一个目的而进化,但结果却经常或通常发现这种进化后的东西具有其它用途。这种情况谈不上什么预谋,这只是一种机会主义”。 笔者不否认粘土作为平台和模子对原始细胞诞生的可能意义,也不否认晶体矿物质在原始地球上最初的小分子有机物产生与累积中的可能作用。但是,生命为何要这样开始呢?怎样在这种粘土环境中诞生出蛋白质、RNA和DNA等生物大分子以及最终过渡到细胞呢?这些都是粘土学说还无法回答的问题。 3. 脂质世界假说(Lipidworld hypothesis) 以色列学者Daniel Segré等提出了所谓脂质世界假说(The lipid world),认为最初的自复制物质是类脂物(lipid-like)(Segré et al. 2001)。在搅拌的情况下,磷脂在水中形成脂质双层结构,在细胞中也是同样的结构,这些分子在早期的地球上并不存在,但其它两性的长链分子也能形成膜。此外,这些脂质体通过脂质分子的插入可能会扩张,在过度扩展的情况下可能会自发地一分为二,子体保存同样的体积和脂质组成。该理论认为,脂质体的分子组成是信息储存的初步方式,进化导致像RNA或DNA这样能有效储存信息的聚合物的出现。 在该假说中,脂质双层膜由各种各样的两性分子化合物混合而成,因而在膜上这些化合物的排列组合方式是巨多的,在这些潜在的组合中,膜上特定的排列应该是有利于艾根的所谓超循环模式,即在膜的位点与捕获在囊膜中的特定化合物之间存在互相催化的正反馈关系。这种位点/化合物对应关系可以传到子代的囊膜,导致能够接受达尔文自然选择的囊膜的不同世系的出现。 笔者也认为,封闭的脂质膜的出现是通向细胞的重要一步,这种结构形成的自发性也为细胞的自然发生奠定了重要的基础,同时,膜的扩张容易导致一分为二也为细胞分裂机制的诞生奠定了一定基础。但是,仅从脂质体的分子组成如何去推导信息储存进而发展到RNA或DNA的出现呢?该理论借用艾根的超循环也是无济于事的。因此,虽然脂质对细胞生命体的出现十分重要,但它还不是最为核心的原因。 4. 多磷酸盐假说(Polyphosphateshypothesis) 美国生物化学家MichaelR. W. Brown和Arthur Kornberg、以及天体生物学和地球化学家Matthew A. Pasek在发表在PNAS的文章中认为,关于生命自然发生的大多数情景面临的一个问题就是氨基酸对多肽的热力学平衡处于个别氨基酸的方向之中,缺失的是驱动多聚化的某种力,而多磷酸盐的特性正好可以解决这一问题(Brown & Kornberg 2004,Pasek 2008)。多磷酸盐由普通的单价磷酸离子PO4−3聚合而成,有几种可能的机制,多磷酸盐引起氨基酸转变为多肽的多聚化,它们也是合成像ATP这样的关键生化物质的合乎逻辑的前体,一个关键的观点似乎是,钙与可溶性磷酸盐反应形成不溶性磷酸钙(磷灰石),因此必须有某种可靠的机制来维持钙离子不引起磷酸盐的沉淀。 笔者认为,多磷酸盐假说关注到最初的生物大分子自发形成的能量基础,不可否认,高能的多磷酸盐是自然形成像ATP这样的能量货币物质的重要基础,且在生命的化学进化具有重要意义,但这也只能是诸多化学进化因素之一。 5. 辅酶世界假说(Coenzymeworld hypothesis) 最近美国遗传学家Alexei A. Sharov和理论生物学Richard Gordon通过将摩尔定律运用于生物进化以及回推法认为,生命起源于97±25亿年前,比地球形成早了数十亿年(Sharov 2006,Sharov & Gordon 2013)。在生物进化中,经验证据表明复杂性每隔3.76亿年翻一倍,就像树木的年龄可用树轮表示一样,生命的年龄也可能用生物复杂性(譬如基因组中功能性非冗余性DNA的长度)来推算。如果将对数转换的复杂性与大进化世系的起源时间作图,得到一直线关系,该直线与X轴相交点(即1个核苷酸)的时间为97±25亿年前。因为太阳系的年龄也只有46亿年,这就是说,生命可能是从更老的星球以某种方式来到地球的。 一些学者基于以下论点对这一假说进行了批评:①由于早期地球缺乏竞争,生物进化速率可能要快得多,②生命起源于几个核苷酸是不可想象的。为了回应这些批评,Sharov提出了一个从类似辅酶分子(功能上与单核苷酸相当)的假想的自然发生情景(Sharov2009,Raffaelli 2011)。笔者认为,这种统计规律性固然有一定的科学价值,但距说明生命起源的机制还相差甚远。 6. 铁—硫世界假说(Iron–sulfur world hypothesis) 德国化学家Wächtershäuser G.于1990年在PNAS上发表了“Evolutionof the first metabolic cycles”一文,提出了一个称之为铁—硫世界的生命起源假说,这也是一种热源起源学说,也与生命起源于海底热液喷口的假说一致。Wächtershäuser(1990)认为,①推动生命起源的能源是铁的硫化物和其它矿物质(如黄铁矿),来自这些金属硫化物的氧化还原反应所释放的能量不仅可用于有机分子的合成,而且可用于低聚物或多聚物的形成,②这样的系统可能进化出成套的能够自我复制的自催化系统以及在我们现在已知的生命形式之前的能够新陈代谢的活的独立实体。Wächtershäuser也是代谢在先论者,认为在生命起源中代谢先于遗传,这里的代谢指一种产能化学反应的循环,而且这种能量能被其它过程所利用,他认为,一旦一种原始的代谢循环建立起来,它就开始生产更复杂的化合物。 笔者认为,这种海底热液喷口假说需要面临的一个问题是,即使这些微生物诞生了,但是,当它们随着混合作用进入茫茫大海时,它们如何能够存活下去?因为这时它们赖以生存的能量来源(大量金属硫化物)已经消失,它们如何能够适应完全不同的生存环境?也许有人会认为,在漫长的地质岁月中这些喷口处的原始细胞中总会有一些个体能够成功演变为适应大海的一般生存环境,但这也只能停留于一种无法证实的假设或猜想之中。在后面还要谈到的利用H2的深海喷口假说也面临同样的问题。 7. 锌世界假说(Zn-World hypothesis) 德国物理化学家ArmenY Mulkidjanian提出了一个称为锌世界的生命起源假说(Mulkidjanian 2009),该学说认为热液中丰富的H2S与原始海洋(或“达尔文水池”)冷水相互作用导致了金属硫化物颗粒的沉降,这就是为何海洋的喷口和其它热液系统分布有带状结构,它也反映了古老的火山成因的块状黄铁矿矿床(VMS)的热液起源。这种结构直径可达数公里,可追溯至太古宙,最丰富的物质为FeS2、CuFeS2和ZnS,还有一些PbS和MnS。ZnS和MnS具有储存辐射(如紫外线)能量的独特能力。在复制分子起源相关的时间窗口,原始大气压力也足够高(> 100 bar)以便在地球表面沉淀ZnS,紫外辐射比现在要强10-100倍,被ZnS媒介的独特的光合作用特性就正好提供了驱动信息和代谢分子合成和耐光的核碱基选择的适当的能量条件。 但是,Mulkidjanian等(2012)注意到,所有细胞的无机物组成都与现代海水的组分不同,他们结合地球化学分析以及现代细胞普遍的无机离子成分需求的谱系分析,重构了最初的细胞的‘孵化场’,推测普遍的原始的蛋白质和功能系统显示出对K+, Zn2+, Mn2+和磷酸盐的亲和性和功能需求。地球化学重建显示,有助于细胞起源的无机离子组成并不存在于今天我们称谓的海底,但与我们今天称谓的内陆地热系统(主要是散发蒸汽的地带)相一致。在缺氧且充满CO2的原始大气条件下,近地热场的水的冷凝与蒸发化学成分将类似于现代细胞的内部环境。因此,进化的前细胞期可能发生在浅的‘达尔文水池’,那里排列着多孔的硅酸盐矿物质,混合着金属硫化物,并富含K+, Zn2+和磷化物。 该学说基于现代生物细胞中的关键离子浓度的比例对首个原始细胞诞生之地进行了推测(充其量只是对‘达尔文水池’假说的扩充),他们认为首个生成的细胞即没有能锁定离子的细胞膜,也没有膜蛋白泵,那么只能与环境保持平衡了,因此现代细胞中的离子构成也许反映了原始细胞起源地的无机离子构成。但是,如何才能保证这一假设是成立的呢? 8. 多环芳烃世界假说(PAH world hypothesis) 一些学者主张生命起源自一些地球大气圈外(星际)的复杂分子,因为通过光谱分析发现,有机分子存在于彗星和陨星之上。美国天文学家Adolf N. Witt 在星系检测出多环芳烃(polycyclic aromatichydrocarbons,PAHs)(Witt 2003),西班牙天体化学家García-Hernández等(2010)在星际检测出PAHs和富勒烯。PAHs是迄今为止在星际空间发现的最复杂的分子。 最近,用斯皮策空间望远镜从环绕一颗正在形成(如同太阳曾经形成的那样)的年轻的恒星HH 46-IR的圆盘里的物质中检测到非常大范围的分子,包括氰化物、烃和CO。2012年,NASA的科学家报道,PAHs在星级介质的条件下发生了转化,通过氢化作用、氧化作用和羟基化作用,变成了更为复杂的有机物,分别是在通往蛋白质和DNA的原材料—氨基酸和核苷酸迈出了一步(Gudipati & Yang 2012)。 笔者认为这一假说一方面提供了星际中存在的有机分子向地球输送的可能,另一方面也指出宇宙空间存在有机分子形成的条件,这或许可以看成是类米勒实验的太空佐证吧,但仅此而已,它无法说明生命起源的具体过程。 9. 代谢在先模型("Metabolismfirst" models) 有若干模型否定“裸基因”("naked-gene")自我复制的观点,而假定原始代谢先出现,这可为后来RNA复制的出现提供环境。在好氧生物为了能量生产的三羧酸循环的中心以及在复杂有机化合物的生物合成中的二氧化碳和氢离子(包括氨基酸和核苷酸)的移动都暗示这是代谢进化的最初部分之一(Lane 2010) 。Smith和Morowitz(2004)认为,如果组分的浓度充足,循环将会自我运转,随着每个中间产物浓度的上升,它就会自发地切换到下一个中间产物。因此,最初的起点不是基因的创造,而只是热力学和化学产物本身。宇宙学家Carroll认为,“生命的目的是为了还原CO2”(代谢在先而不是遗传在先情景的一部分)(Musser 2011)。 笔者认为,这一理论虽然还十分粗糙,但是提出了有价值的观点,即生命不可能起源于所谓的“裸基因”,至于“生命的目的是为了还原CO2”这样的说法,虽然没有揭示生命的本质,但至少比无目的性要强。 10. 自催化假说(Autocatalysis hypothesis) 美国理论生物学家、复杂理论的领军人物Stuart A.Kauffman在“The Origins ofOrder: Self-Organization and Selection in Evolution”一书中提出生命最初起源于自催化化学反应网络(Kauffman 1993)。所谓自催化指能催化自我形成的物质,因此也具有简单的分子复制者的特性。Rebek等构建了氨基腺苷(amino adenosine)和五氟苯酯(pentafluorophenyl ester)与自催化剂氨基腺苷三酸酯(AATE)的混合系统,其中一个来自实验的系统包含了多种能自行催化自身合成的AATE。此实验证明了自催化可能在一个分子种群的不同遗传实体间进行竞争的可能性,这可解释为一种初步的自然选择方式(Dawkins 2004)。 依笔者之见,这与艾根的以分子自组织为基础的超循环理论大同小异,没有本质的差别;此外,没有隔离(细胞膜)的系统,所谓初步的自然选择也是不可能发生的。 11. 自组织和自复制假说(Self-organizationand replication hypothesis) 自组织和自复制常被认为是生命系统的标志,但是也有非生物分子在合适的条件下呈现出类似特性的许多例子。譬如,美国植物学家William Martin和Michael J. Russell认为,从环境中的细胞膜的物理分隔与自我支撑的氧化还原反应的自组织是生物最保守的特性,因此,具有这种特性的无机物就最有可能是生命最后的共同祖先(Martin & Russell 2003)。后来,Palasek(2013)指出,由于热液喷口的物理因子,RNA分子能自发地进行自我组装。在宿主体内病毒的自我装配对生命起源的研究也有借鉴意义,因为它使生命能起始于自我组装的有机分子的理论更为可信(Nussinov et al. 1997,Vlassov et al. 2005,Koonin et al. 2006)。 笔者认为,该学说指出细胞膜的物理隔离以及自我支撑的氧化还原反应是生命最后的共同祖先的特性这一点其实是对判断原始细胞标准的一种说明。至于以病毒为例的RNA的自我装配在生命起源早期的作用则难以确认,因为,对现代生命来说,没有宿主,病毒也将不复存在。 美国法学教授、反达尔文主义者詹腓力(Phillip E.Johnson,1940-)在Darwin On Trial(译文:审判达尔文)一书中批评道,“既然在实验上取得成果的前途不佳,有些科学家干脆就从计算机模拟方式下手,以免碰到实验性的障碍……一般科学家已经假定生命是自生自发而来,以后不断进化到更复杂的结构。果真如此,那么万物当然需要有自发性的自我组合能力了……运用这种假说,科学家可以设计一些电脑模式来模拟生命起源及其进化。至于这个模式与现实有何关联,倒完全是另一个问题了”(Johnson 1991)。 12. 深部热液生物圈模型("deep-hot biosphere"model)
20世纪70年代,美籍奥地利裔天体物理学家Thomas Gold(1920 – 2004))提出了深部热液生物圈模型,认为生命最初不是起源自地表,而是在地表的数公里之下(Gold 1992,2001)。20世纪90年代后期在深部岩石中发现纳米微生物(比细菌更小的丝状结构,但可能含有DNA)可能看作是对Gold理论的支持。 现在已经熟知,从地表到之下5公里都存在丰富的微生物,主要是嗜极的古细菌而不是真细菌,后者更多地分布在一些容易接触的环境之中。Gold声称,从深不可及之处食物的涓涓细流对生存来说是需要的,因为在一个水坑中诞生的生物总会将其中的有机物质消费一空,最后自己也会灭绝。Gold的理论认为,这样的食物流源于地幔中原始甲烷气的溢出,深部微生物(远离沉积碳化合物)食物供给更通常的解释是有机体靠水与岩石中的(还原性)铁化合物之间相互作用释放的氢气来维持生活。 笔者认为,深部热液存在微生物是事实,但到底那里是否是生命诞生之地又是另外一回事,既不能排除这种可能,也无法确认亦如此。但是,这些假想的共同祖先又如何能从深达数公里的地下顺利到达地面并成功转变自己的代谢系统以适应新的缺乏甲烷或H2的地表呢?如果地表有这样的能量物质,那它们又何必隐匿于地下而被创造呢? 13. 深海喷口假说(Deep sea vent hypothesis) 深海喷口或碱性热液喷口假设认为,生命可能起源于海底的热液喷口,在那里富含氢气的液体从海底涌出(如式8-1),在这样一个系统中,持续的化学能由氧化还原作用所提供,在那里,电子供体(如H2)与电子受体(如CO2)发生反应。 3Fe2SiO4+ 2H2O → 2Fe3O4 + 3SiO2 + 2H2 (8-1) Martin和Russell(2003)认为,碱性喷口创造了一个自发的质子动力势化学渗透梯度,这种条件是生命自然孵化的理想场所。这里的细微隔室,为有机分子的浓缩提供了一种自然的方式,如Wächtershäuser设想的那样,这些由铁—硫矿物(如四方铁硫矿)构成的隔室天生就有催化特性(Lane 2010)。离子的跨膜运动源于以下两个因子的组合: ① 由浓度梯度引起的扩散力—包括离子在内的所有颗粒物倾向于从高浓度向低浓度扩散; ② 由电势梯度引起的静电力—像质子H+这样的阳离子倾向于沿电势向下扩散,而阴离子则方向相反。 这两个梯度合在一起就是电化学梯度,为自发的合成提供能量。质子动力势也能表述为储存为跨膜(质子浓度和电势不同)的质子与电压梯度相组合的势能的一个度量。 笔者认为,这里只是描述了现代化能自养菌的特性,此外,这一假设面临与前面说过的这些假想的共同祖先在被稀释到茫茫大海中之后如何能够成功适应完全不同的新的环境的问题。 14. 热合成世界假说(Thermosynthesisworld hypothesis) 现今发酵的生物能量学过程通过三羧酸循環或乙酰辅酶A途径来进行,两者都与原始的铁—硫假说相联系。而荷兰的生物化学与物理学家Anthonie Muller提出的热合成假说则考虑化学渗透的生物能量学过程,这在细胞呼吸和光合作用中扮演着重要角色,比发酵更为基础(Muller 1985,1995)。在热合成世界,维持化学渗透的ATP合成酶被认为是在现存的酶中最接近最初的代谢过程的。 首先,生命需要能源去推动凝聚反应以产生出蛋白质的多肽键和RNA的磷酸二酯键。一般来说,关于今天的ATP合成酶的结合变化机制的热量变化,“第一个蛋白质”应该结合了底物(多肽、磷酸、核苷酸、RNA‘单体’)并将他们浓缩成一个反应产物,这个产物会保持结合直到经历一个温度变化它才被热诱导去折叠所释放。热合成世界的能源来自热循环,就是一个对流的水流中原始细胞悬浮的结果,正如一个火山温泉的情形一样,对流解释任一生命起源模型所需要的自组织和耗散结构。热循环在萌发和细胞分裂过程中仍然扮演着普遍角色被认为是原始热合成的遗物。 通过磷酸化细胞膜脂质,这个‘最初的蛋白’使含有蛋白的脂质原始细胞具有了选择优势。一开始,这个最初的蛋白也合成了一系列蛋白,其中只有极少数具有热合成能力。正如美籍英裔数学物理学家Freeman Dyson在“Origins of Life”一书(Dyson 1985)中所设想的,这个最初的蛋白功能性地繁殖了:它制造了相似能力的后代,但它没有拷贝自身。功能后代由不同的氨基酸序列组成。假设在随机合成的RNA中,RNA序列根据最初蛋白合成的速度和效率增加来选择,譬如根据作为信使RNA、转移RNA或核糖体RNA的生成,或者更一般地,RNA世界的所有成员被生成与选择。这样,在理论上热合成世界可解释遗传体系的起源。 铁—硫世界确认了一个最简单的循环路径,因此假定了酶的存在,但是,热合成世界甚至没提及一个通路,也没有假定通常酶的存在:ATP合成酶的结合变化机制类似于一个产生自由能的物理吸附过程(Muller &Schulze-Makuch 2006),而不是一个正常的减少自由能的酶的机制。RNA世界也意味了若干酶的存在。有人声称,靠随机来产生一个酶,这是绝不可能的事(Orgel 1987)。 这是一个蛋白质在先的假设,但笔者认为它在如何通过蛋白生成DAN和RNA的说法上并不能令人信服。 15. 放射性海滩假说(Radioactivebeach hypothesis) 研究天体生物学的美国学者Zachary Adam提出了一种称为放射性海滩的生命起源假说(Adam 2006)。他认为,曾经在某一时候发生过的潮汐过程(当月亮非常接近时,可能在原始海滨的高水位标志区浓缩了铀和其它放射性元素颗粒)可能导致了生命构建物质的起源。这样的放射性海滨可能为有机分子(氨基酸、糖)的产生提供了足够的能量,放射性的磷铈镧矿(monazite)还可释放生物易获得的可溶性磷。这样氨基酸、糖和溶解性磷化物就同时形成了。在这种反应中所遗留下来的一定浓度的发射性锕类可能形成了有机金属复合物的部分,这些复合物可能是生命过程的早期催化剂。 笔者认为这一假说为原始生命诞生之地提供了一种可能,它假设放射性物质为从无机化合物合成简单有机化合物提供了能量,并假想了早期的催化剂,但是,生物大分子以及最后的细胞又是如何起源的呢? 16. 紫外线和温度辅助的复制模型(Ultraviolet and temperature-assisted replication model) 紫外线和温度辅助的复制模型主要是基于生命起源的热动力学视角。墨西哥研究核物理学(Nuclear Physics)和进化物理学(Physics of Evolution)的学者Karo Michaelian 认为,任何关于生命起源的模型都必须考虑这样的事实,即生命是一个不可逆的热动力学过程,它的产生与持续归因于它产生熵,熵的产生对生命来说并不是偶然的,而是生命存在的根本原因。现在的生命通过促进基于蒸散的水循环而增加地球的熵的生产;因此,如果现今生命的热力学功能是伴随水循环而产生熵,那在生命诞生之初也应该如此;事实上,水溶液中的RNA和DNA是200-300 nm波长范围的紫外光的非常强烈的吸收者,这是能穿过浓厚的前生物大气层到达地表的太阳光的一部分(Michaelian 2009,2011a)。 Cnossen等(2007)认为,在太古宙时期到达地球表面的波长在260 nm处(也是RNA和DNA吸收最强烈的波长)的紫外光的量能是现在的31个数量级。紫外光在太古宙海面被有机分子的吸收和耗散显著增加了表层温度,导致蒸发增加,因此也增加了原始的水循环。因为光能量光子的吸收和耗散是一种熵的生产过程,Michaelian认为热动力学过程应该促进了利用紫外光进行的RNA和DNA的非平衡的自然合成过程(Powner et al. 2009)。 有一个不必借助酶就可以解释RNA和DNA复制的简单机制,它可由这同样的热力学框架所提供,假设生命出现时原始海洋的温度已经冷却到了差不多RNA或DNA的变性温度之下(根据南非巴伯顿大约35—33亿年前的绿岩带燧石中的18O/16O的比率,表面温度估计约为70±15 °C,接近RNA或DNA的变性温度)(Lowe & Tice 2004)。在晚上,表层水温将降低到变性温度以下时,单链RNA/DNA可做为模板形成双链的RNA/DNA;到了白天,RNA和DNA将吸收紫外线,将其转化为热而使局域温度升高到可使RNA和DNA变性,这种拷贝过程在昼夜循环中反复进行(Michaelian 2010,2011b)。这种温度辅助的拷贝机制与实验室日常用于扩增DNA片段的PCR(聚合酶链式反应)相似。Michaelian认为,传统的关于生命起源的研究期待去描述近于平衡条件的生命诞生,但这是错误的,必须考虑非平衡的条件,特别是熵生产对生命诞生的重要性。 笔者认为该学说提供了一个新奇的视角,但是,问题是这些RNA和DNA最初是怎样或为何产生的呢? 17. 水泡假设(Bubbleshypothesis) 冲击海岸的波浪产生精致的泡沫,席卷海洋的气流具有驱动表面颗粒冲上海岸的倾向,这种海岸线浮沫与被风吹拂的有机颗粒可在海滩相互作用,而滨海浅滩上的海水会更加温暖,可能通过更快的蒸发浓缩有机分子;主要由水构成的水泡很快就会破裂,而含有两亲分子的水泡就会稳定得多,允许其中的关键反应有更长的时间进行。两亲分子具有亲水的头部和疏水的尾部,一些两亲分子在水中有自发形成膜的倾向,一个球形含水的封闭的膜就是现代细胞膜的假想的前体。如果一个蛋白能增加亲体水泡的完整性,这个水泡就会有优势,会被置于更接近于自然选择等待名录中的顶端。当水泡破裂时可想像为原始的繁殖,同时向周围的环境之中释放“实验”的结果。一旦足够 “正确的构建原料”被释放进环境,就可能出现最初的原核生物以及真核生物和多细胞生物的发展(Panno 2005)。 英国物理化学家凯恩斯·斯密斯(1995)提出了这样的一种设想:“在缺氧状态下,海洋分子将‘生命分子’积聚起来。海洋就像一个盛着营养汤的巨大的碗,为‘生命分子’的积聚提供了场所,其它则由机会来完成。在这只大碗中,各种分子不断结合、破裂。有些分子之间的结合会比其它分子的结合更为稳定,于是,这种结合就形成了汤中凝聚的小飞沫或小凝块。海浪时常将这些不断增大的飞沫打散成小块,这些小块飞沫也同样吸收了新的物质。最后,这种分子结合体中有一个—也只需要一个即可—终于爬上了达尔文描述的进化的阶梯。它可以繁衍,可以将其特征传给后代”。 笔者认为,水泡假设提供了原始细胞的一种可能模式,与脂质世界假说或“原始汤”假说或多或少有些相似,但同样未深入涉及生物大分子的形成问题。 18. 多起源假说(Multiplegenesis) 在地球历史的初期,不同形式的生命可能准同步(quasi-simultaneously)出现(Davies 2007)。其它的形式可能绝迹了,由于其不同的生物化学过程(譬如利用砷代替硫)而留下了特别的化石,或作为嗜极生物而幸存下来,或者简单地仅仅是由于与现在的生命树上的生物相似而未被注意。Hartman(1998)通过许多理论的整合提出了以下观点:最初的生物是自我复制的富含铁的粘土,它固定CO2并将其转化成草酸或其它二羧基酸,接着,这个由能复制的粘土和它们的代谢表型构成的系统进化到需要固氮能力的富含硫的温泉区域。最后,硫被整合到一种能合成核苷酸和磷脂的进化系统之中。如果生物合成重述了无生源说,那么氨基酸的合成先行于嘌呤和嘧啶碱基的合成。此外,氨基酸硫酯聚合成多肽先于通过多聚核苷酸的氨基酸酯的直接聚合。 已故的美国生物学家LynnMargulis(1938–2011)的内共生理论认为,不同形式的古细菌通过共生形成了真核细胞(Margulis & Sagan 2001),在不同古细菌之间遗传物质的水平转移促进了这样的共生关系,因此,许多不同的有机体可能参与贡献了普遍称之为我们现代生物的共同祖先(LUCA)的构建。 19. 对生命起源依旧迷惘 苏联学者奥巴林在他1924年出版的《生命的起源》中说,“很遗憾,细胞的起源仍然是个问题,也是全部进化理论中最黑暗的一点”;1988年,德国学者道斯(Dose K)在他的一篇论文中说,“在化学和分子进化领域里,关于生命起源的实验已做了30多年;这些试验实际上扩大了这个问题,而没能找到解决问题的办法。目前,所有关于这些理论的讨论和实验,不是陷入僵局就是对无知招供了”;1998年,美国学者巴达(Bada J)在他的一篇文章中说,“今天,当我们告别20世纪时,我们仍然面对进入20世纪时的最大的未能解决的问题:生命怎样在地球上开始?”(雅荷雅2003)。 美国生物化学家贝希(Michael J. Behe)在“Darwin’sBlack Box: The Biochemical Challenge to Evolution”(译文:达尔文的黑匣子—生化理论对进化论的挑战)中说,“当化学家努力用实验和计算来验证生命到底从何而来的时候,进化生物学家们却没有做出任何努力通过实验和计算验证分子层次上的进化情景。结果,进化生物学就停留在50年代早期支配生命起源研究的相同的思维框架中”(Behe 1996)。我倒觉得贝希这样的批评不无道理。 美国法学教授、反达尔文主义者詹腓力(1999)说,“能够想象或构想一个理论,与可能用实验证明出来,还有天壤之别……没有实验证据支持任何一种理论,反而为无限制的臆测打开了大门”。他对令人眼花缭乱的各种各样的生命起源理论的这种批评也不是没有一点道理。 我很赞同法国哲学家(也是神学家和古生物学家)德日进的看法:“‘有机化合物’的世界才是我们的……虽然它形式十分的多,比矿物不知多过凡几,但因为只构成地球物质的很小部分,以致地球化学只视它为旁支……有机物必须是看作在地球初生时就已开始,一点也不比无机物为迟……若不在开始时就真的存在,则它们以后永不会发生……为什么要在渺茫的太空中寻找地球的起源呢—这岂不是不可理喻吗?早期的地球,它本身所有的化学组成,这本身整个乃就是我们梦寐以求的生命来源。当它生成时,它已先天地(如果我能这么说的话)带来了‘前生命’了,而且是带有‘一定量’的”(de Chardin 1955),当然,这里的有机物应该限定于像氨基酸、碱基(如嘌呤)、单糖类、有机酸等生命的基本构件物质。 结语 在过去的近一个世纪中,关于生命起源的追求从未停息,争议也从未销歇。生命起源这个领域就像一个熙熙攘攘的集市,各种学说飘来荡去,恰似西风中坠落四散的黄叶,可而今这个神秘的“真容”依然还深藏于灰雾濛濛之中。在这里,罕见的是禁锢,常见的是草率仓猝,几乎人人可以自炫一番,个个为自己的空想得意洋洋,因为无法证实,也不能证伪。但大家迷茫依旧,因不知路有多远,山有多遥。借引歌德在“你为何给我们深邃的眼力”一诗(创作于1776年)中的一句名言:“一个空洞的梦使他无事忙碌也是幸福,一种想象使他内心感到充实也是幸福”。历史舞台上的生命起源论,莫不亦如此? 我们是否应该重温德国伟大哲学家黑格尔(Georg WilhelmFriedrich Hegel,1770年-1831年)的哲言:“有机的东西是现实的东西,这种东西自己维持自己,经过它自身内的过程;它是它自己的普遍东西,这种普遍东西把自己分裂为自己的各个部分。这些部分在创造整体时,就扬弃了自己”? 我们是否应该邃思法国哲学家德日进(Pierre Teilhardde Chardin,1881年-1955年)智语:“一向被视为宇宙结构之点来处理的个样事物,现在都可以看做是一种无限定的时间纤维中的瞬刻了。张眼望之,物质各元素继往开来、延亘不绝,都是那根植于无量度的过去并发展向遥远渺不可及的未来之系列中的一部分罢了”。在探索的茫茫黑夜之中,如何找到一盏照耀的明灯? 主要参考: 谢平. 2014. 生命的起源—进化理论之扬弃与革新.北京:科学出版社(英文:Xie P. 2014. The Aufhebung and Breakthrough of the Theories on the Origin and Evolution of Life. Beijing: Science Press) 作者:谢平 来源:科学网 人类的迷惘:生命源自何方? | 








 提升卡
提升卡 置顶卡
置顶卡 沉默卡
沉默卡 喧嚣卡
喧嚣卡 变色卡
变色卡 千斤顶
千斤顶 照妖镜
照妖镜