|
���ڱ�Ƥ���ʸ��ǣ������½��ֲ�����۳����ʺͰ�˪״��������ʧ���ܻᵼ�¹���ı��ͣ�����Ҷ���Ǹ���������״����ˣ�������Ƥ����ȱ����ֲ�ﲢ�������Ʊ�Ƥ��������ϳɵĵ��ػ��ƽ�������ũ�������������ʡũҵ��ѧԺ���о���&֣�ݴ�ѧũѧԺ�ڡ�Theoretical and Applied Genetics����5.699/Q1���������¡�BrWAX2 plays an essential role in cuticular wax biosynthesis in Chinese cabbage (Brassica rapa L. ssp. pekinensis)�����������ڸ�ȫ����˽�ֲ����ʲ�����л���磬�������ڼ�����������������ɫ�������Ͳ�Ʒ�֡� ��άɭ�ṩ��ת¼�����������
��������������ֲ�����Ľ��ʲ����ڱ���ֲ����������ͷ�����в�ȷ������Ÿ��ָ��������á�����ͻ���徥Ҷ������й����ڷḻ��ײ�������Դ��ѡ������Ʒ�ַ��������Ҫ���á�Ȼ�����йش�ײ˹�����״�Ļ���ȴ���б����� ��������ֲ����ϣ� �ױ�������˫������ƷϵR16-11��P1�������Խ���ײ�ƷϵY1211-1��P2�� �Ӵ���F1��F2��BC1P1��BC1P2��BC1P1��BC1P2�ֱ���F1��R16-11��Y1211-1�ؽ���ã� 18���й����̦ܿ���ϣ�11�ֲ��ĺ�7�ִ�ײˣ���10�����ʴ�ײ˲��� ������ RNA-seq��BSA-seq��qRT-PCR
����������1�� Y1211-1�Ĺ�����״���ɵ��������Ի������ R16-11�����������ٱ��棬�绨��ͼ1a��������Ҷ��ͼ1b�������ӡ���ѿ�Ⱦ��������ǡ����֮�£�Y1211-1��Ӧ���ٵ���ۣ�ͼ1d��e���������ڹ⻬����ɫ���õ���ɨ��羵��Cryo-SEM������˻���ҶƬ�����ܶȣ�����R16-11��ҶƬ���渲���Ŵ�����������ҪΪ��״�Ͱ�״��ͼ1c������Y1211-1��ҶƬ���������������ǣ�ͼ1f����˵��Y1211-1�Ĺ⻬��������������������ġ�
ͼ1 �����ױ���R16-11��Y1211-1����Ƥ���ʵı�������
��F1Ⱥ���У����и��嶼���ֳ����ʱ��ͣ�R16-11Ҳ����ˡ�F2��BC1P2Ⱥ����ڱ��ͷ�������F2СȺ�����ʺ���������ֱ�Ϊ148����52���������Ϊ3:1��F2��Ⱥ��ķ����ҲΪ3:1��BC1P2��Ⱥ�����ʺ��й�����������ֱ�Ϊ526��494������Ϊ1:1��200��BC1P1ֲ����۾�����״����Щ���������Y1211-1�Ĺ����������һ�����������Ի�����Ƶģ���1�������ǽ���λ������ΪBrWAX2��
��1 R16-11��Y1211-1�ӽ����������״���Ŵ����� ��2�����й����ֲ���У���Ƥ���IJ����ᷢ���ı� Ϊ�о�����ı����Ƿ����������Լ����йأ�ͨ��GC-MS�����W-bulk��G-bulkҶ�ӵı�Ƥ���ĺ�������ɡ�������ֲ����ȣ�����ҶƬ�ϵ�������������46.6%��G-bulk��������ȩ���ܺ����ֱ��½���79.7%��95.3%��G-bulk�е�ȩ���ܺ�����Լ��W-bulk�е�2.86����C29���������飩��C30ȩ����ʮ�飩���ֱ�ռW-bulk��G-bulk����������Լ59.2%��55.6%��Ȼ��G-bulk��C29��������������83.7%��C30ȩ����������Լ3.4�������������Y1211-1���й���ı������������������٣���������������������ɵġ�
ͼ2 W-bulk��G-bulkҶƬ�еĽ��ʲ����ɷ�
��3��BrWAX2����ľ�ϸ��λ Ϊ�˳�����λBrWAX2����W-pool��G-pool����BSA-Seq��������Ⱦɫ��A09����95%������ˮƽʶ���һ����37.35��38.88 Mb��1.53 Mb��ѡ����W-pool��ƽ��SNPָ��Ϊ0.60������Ӧ����G-pool��ƽ��SNPָ��Ϊ0.11������������û������������һ������������Y1211-1�Ĺ�����״��һ�����Ժ˻�����Ƶļ���һ�¡� ����BSA-Seq�������ں�ѡ��������30��KASP��ǣ�����ɸѡ�����ױ���12��KASP��DZ��ֳ����õĶ�̬�ԡ�BrWAX2λ����LF2-K24��LF2-K17���Ŵ�����ֱ�Ϊ0.5��0.1 cM���Ŵ�ͼ���б�ǵ�˳��������ͼ���е�˳��һ�£�ͼ3a���� ʹ��flanking��ǣ�LF2-K8��LF2-K30��ɸѡ��967���й����F2ֲ���������90�������壨ͼ3b�������LF2-K30��BrWAX2�����룬����90��������������LF2-K8ɸѡ������BrWAX2�������ڱ��LF2-K36��Ⱦɫ��A09ĩ��֮���100.78 kb�����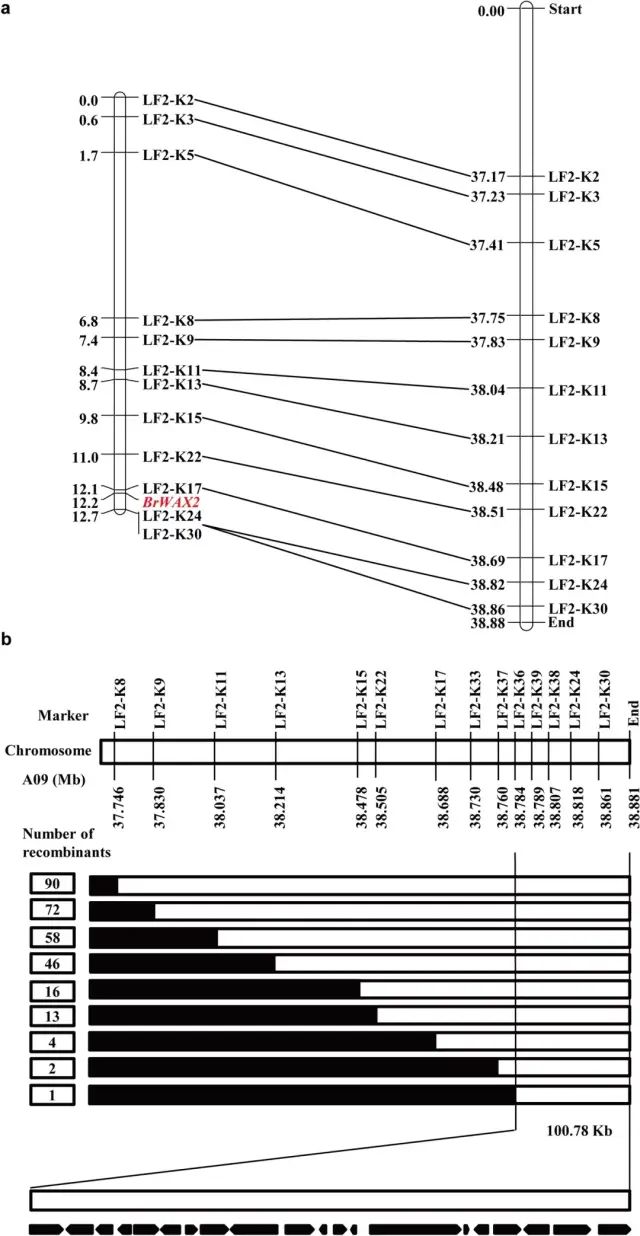 ͼ3 ��ײ�BrWAX2����ij����;�ϸ��λ
��4����ѡ������� ����BrWAX2�ľ�ϸ��λ�������Brassica���ݿ��з�����100.78kb����ڵ�DNA���У��������Ͻ�����˱Ƚϻ���ע�͡������ʾ������ͼ����ʶ���20��ע�ͻ�Ԥ�����2����������BRAD����20�������ע�ͷ���������ֻ��һ����ΪBra032670�Ļ�������Ǻ�ѡ���û��������Ͻ��е�CER1ͬԴ��CER1����һ��ȩ����ø���ڽ��ʲ�������ϳɹ����д�C30ȩת��ΪC29������
��2 BrWAX2λ���ѡ�����ע�ͻ���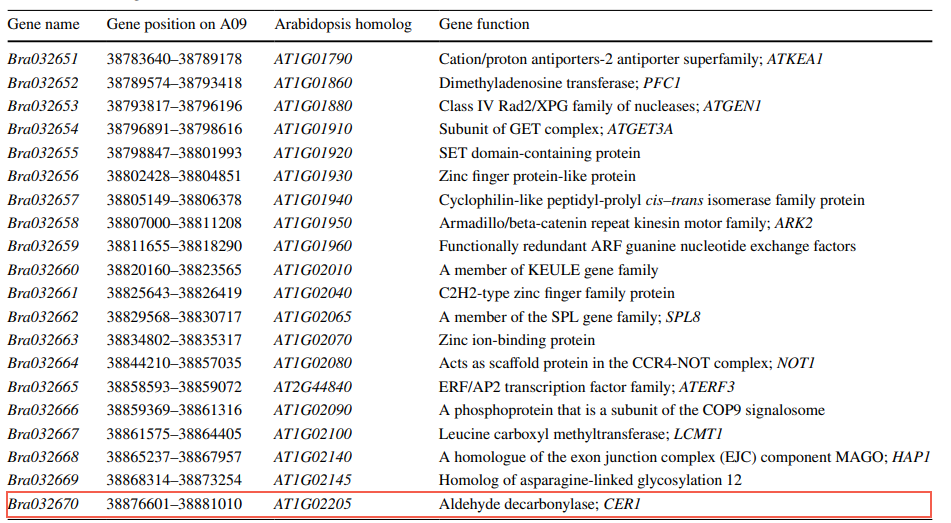
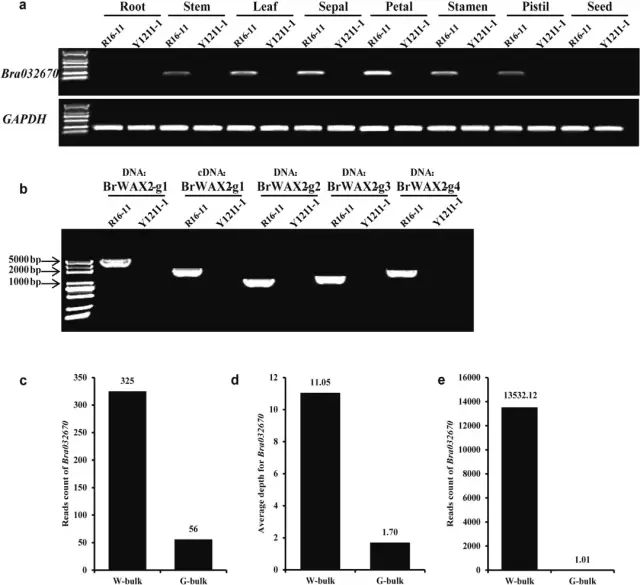
ͨ���붨��RT-PCR��qRT-PCR�����Bra032670�������ױ���ͬ��֯�еı��������Bra032670��Y1211-1�еİ�����֯�о�δת¼��ͼ4a�������֮�£���R16-11�еľ���Ҷ����Ƭ�����ꡢ����ʹ������Ҫ�ߵöࡣRNA-Seq������ʾBra032670�Dz����������Ļ������ڹ⻬���еı���ˮƽԼΪ������13000����ͼ4e�����������Bra032670������BrWAX2����ܵĺ�ѡ���� Ϊ�˼�����ѡ�������ױ�ϵ�е����У�ʹ������BrWAX2-g1��Bra032670�Ļ��������кͱ������н��������Ͳ���ʹ��gDNA��cDNA��Ϊģ���Y1211-1��û�л���������ͼ4b�����Ʋ���Y1211-1��ȱ��Bra032670����W-bulk�бȶԵ�Bra032670�����BSA-Seq����������Ϊ325����G-bulk��ֻ��56����ͼ4c����Bra032670��ƽ��BSA-Seq�����W-bulk��Ϊ11.05������G-bulk�н�Ϊ1.70��ͼ4d����3��G-bulk�ظ��У�Bra032670��RNA-Seq������Ϊ1.01�����֮�£�Bra032670������W-bulk��ƽ������������ߴ�13,532.12��ͼ4e������Щ��ͨ���������ݽ�һ��������Bra032670������Y1211-1��ȱʧ��
ͼ4 BrWAX2��ѡ������� ��5�����ʺ���ת¼����� ��W-bulk��G-bulk֮�乲��1399��DEGs��KEGG�����������������ʡ�ľ˨�ʺ���������ϳɡ�������л�������ϳɺʹ�л;��������������;����ͼ5a����������Щ;���еĴ����������G-bulk�еı���ˮƽ�ϵͣ������ֲ�����������Ľ���һ�¡� ��W-bulk��ȣ�G-bulk�в���������ϳɵ������γ�;���Ļ�����CER1��Bra032670����CER3��Bra006802����MAH1��Bra027906�������µ������������γ�;����G-bulk���ܵ����ơ�������ʲ���ת�˵Ļ���ABCG12��Bra031249����G-bulk���ϵ���������ڽ��ʺ�������ϳɵĻ���WIN1��Bra026140���ϵ�����MYB96��Bra035893����MYB96��Bra010085����G-bulk���µ������⣬�����������ϳɵĻ�������CYP77A6��Bra029852����CYP86A4��Bra032642����HTH��Bra003862������G-bulk�������ϵ���ͼ5b������һ��������л��ػ������qRT-PCR��������RNA-Seq���һ����ͼ6����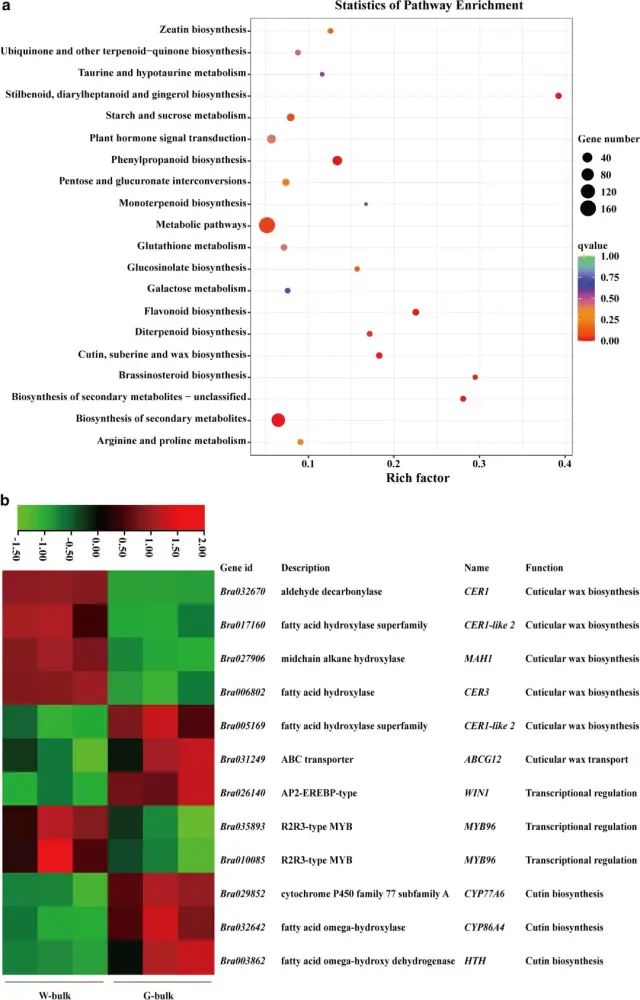 ͼ5 ���ʺ⻬����ת¼�����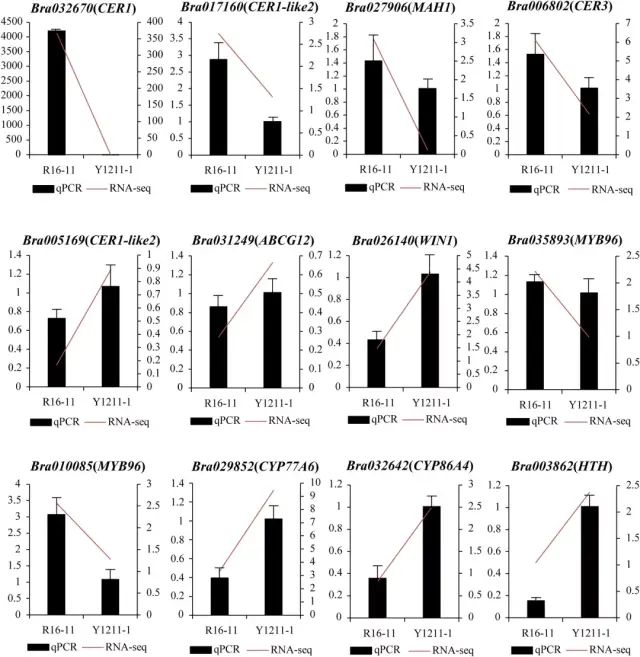 ͼ6 ��qRT-PCR��֤RNA-Seq���ݵı��� �����������о�ȷ���˴�ײ˱�Ƥ������ϳɵ�BrWAX2������BSA-Seq��ͨ��KASP����������������BrWAX2����λ��100.78 kb�����䡣RNA-seq����ע�ͷ�����������������б����������������ȩ����ø��Bra032670��BrWAX2����ܵĺ�ѡ����BrWAX2��ȱʧ���Լ������γ�;���������������Ľ��ͣ������������������������˹�����͡�
ת¼���ʾ��ײ˱�Ƥ���ʵ�����ϳɻ��� |  ¥��|
������ 2022-6-8 10:18
¥��|
������ 2022-6-8 10:18
 ������
������ �ö���
�ö��� ��Ĭ��
��Ĭ�� ������
������ ��ɫ��
��ɫ�� ǧ�ﶥ
ǧ�ﶥ ������
������